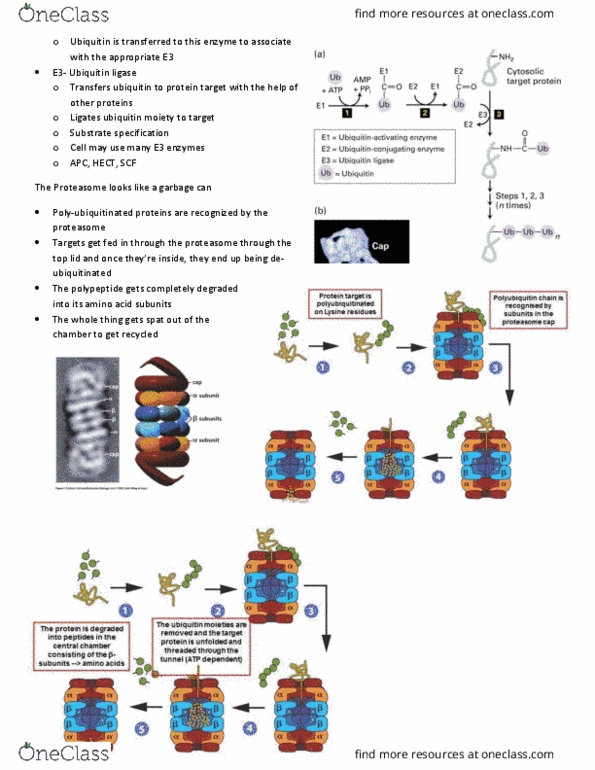
BIOL 200 Lecture Notes - Nuclear Pore, Lysine, Dosage Compensation

6 Apr 2012
School
Department
Course
Professor

Document Summary
As we have learned before, dna does not exist naked in the cell. It undergoes extensive wrapping to form chromatin in the mitotic phase. Through associations with histones 1-4, dna is wrapped into higher order nucleosomes. These nucleosomes can undergo further folding into 30nm fibers, 300nm fibers, 700nm fibers and eventually 1400nm chromatin fibers. While dna is normally found in its wrapped up form, it must be unwound in order for protein factors involved in transcription to bind to the dna and carry out transcription. Modification in chromatin structure severely affects gene expression (transcription) levels. Unwinding of the chromatin facilitates binding of transcription factors thus increasing the rate of transcription. This unwinding can be carried out by a series of proteins and complexes, including histone modifying enzymes, chromatin remodelling complexes and components of rna polymerase ii. Discovery of chromatin unwinding began with work on saccharomyces cerevisiae (baker"s yeast)