🏷️ LIMITED TIME OFFER: GET 20% OFF GRADE+ YEARLY SUBSCRIPTION →
Pricing
Log in
Sign up
Home
Homework Help
Study Guides
Class Notes
Textbook Notes
Textbook Solutions
Booster Classes
Blog
Home
Class Notes
1,200,000
CA
670,000
BIOL 201 Lecture Notes - Chloroplast, Signal Recognition Particle, Rubisco
33
views
19
pages
orangeyak693
27 Feb 2014
School
McGill University
Department
Biology
Course
BIOL 201
Professor
Greg Brown
Like
For unlimited access to Class Notes, a
Class+
subscription is required.
Get access
Yearly
Monthly
Yearly
Grade+
20% off
$8
USD/m
$10 USD/m
Billed $96 USD annually
Homework Help
Study Guides
Textbook Solutions
Class Notes
Textbook Notes
Booster Class
40 Verified Answers
Class+
$8
USD/m
Billed $96 USD annually
Homework Help
Study Guides
Textbook Solutions
Class Notes
Textbook Notes
Booster Class
30 Verified Answers
Continue
Related Documents
BIOL 201 Lecture Notes - Lecture 16: Amphiphile, Membrane Protein, Mitochondrion
crimsonjackal99
BIOL 201 Lecture Notes - Lecture 20: Translocon, Ldl Receptor, Alpha Helix
blackhedgehog334
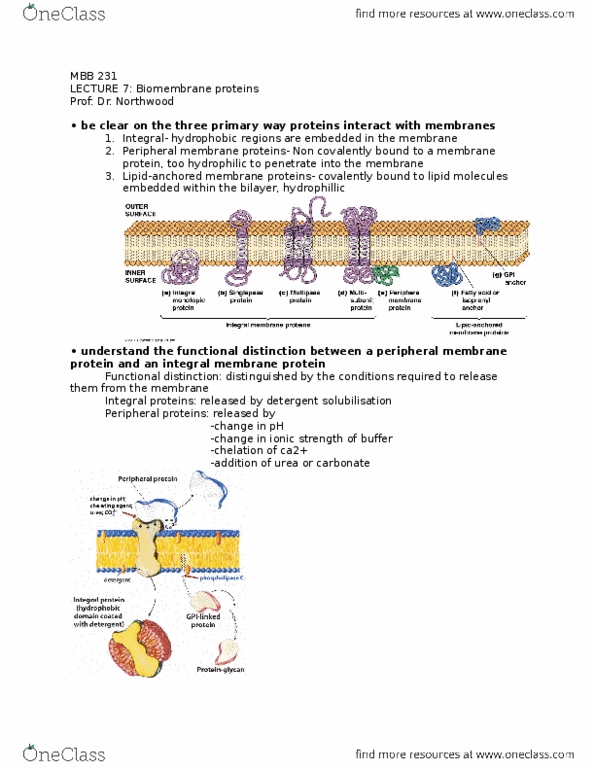
MBB 231 Lecture Notes - Lecture 7: Integral Membrane Protein, Peripheral Membrane Protein, Isoprene
coralgrasshopper248