BIOL 201 Lecture Notes - Profilin, Listeria Monocytogenes, Fimbrin

8 May 2012
School
Department
Course
Professor
Document Summary
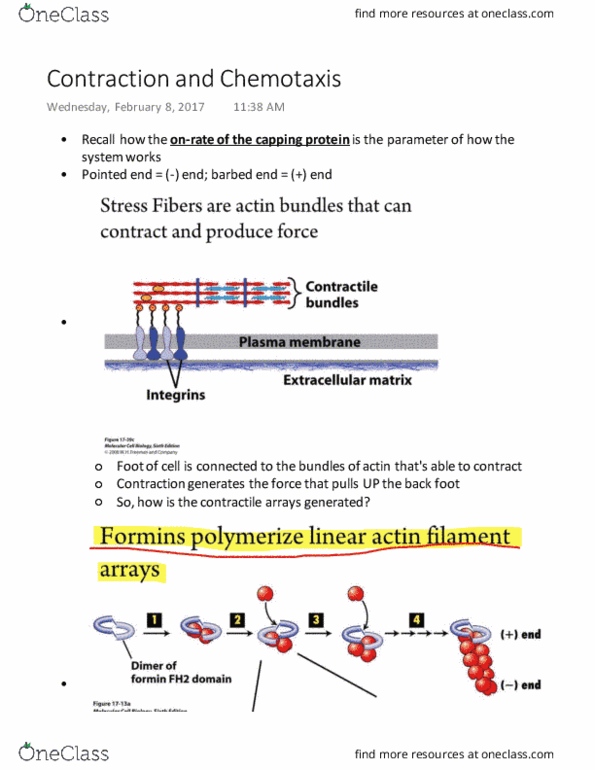
Plus-end actin binds to the end with an on-rate constant (multiplying concentration to the constant) Balance of two numbers gives a critical concentration (below - plus end will shrink) Atp hydrolysis cycle that happens at the same time, actin polymerizes in atp state. Phosphate eventually dissociates to be left with adp-actin. C+c means that actin filaments will depart from minus end but adding to the plus end. The net length of filament not changing - treadmilling, but filament translocates in space. Should be able to calculate conc. at which this process occurs. Adp-actin - actin has to regenerate into atp state before it can repolymerize. The number of proteins that the cells have created allows them to modify the underlying properties of the actin network. Formins sit at plus end and control the rate of which the actin filament grows. Other formins gate the polymerization and slow down the filament elongation.