BIOC 212 Lecture Notes - Lecture 15: Protocadherin, Rac1, Cadherin

22 Aug 2012
School
Department
Course
Professor
Document Summary
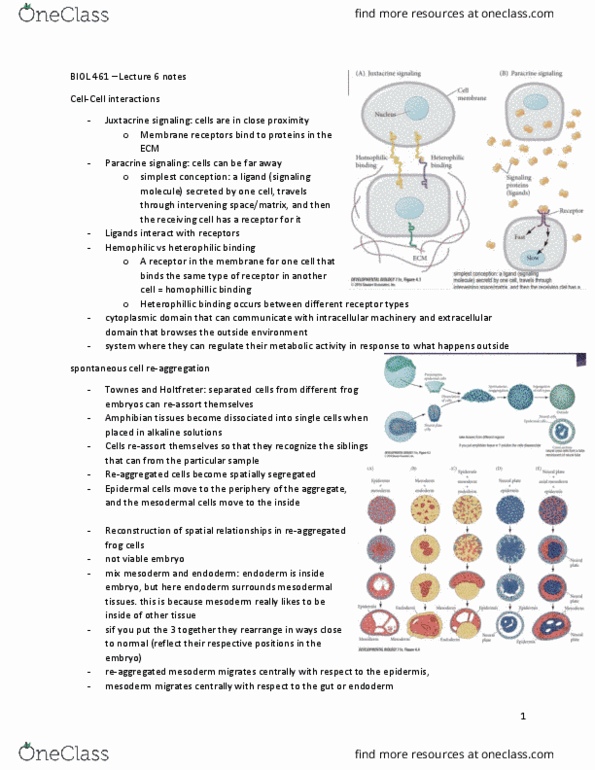
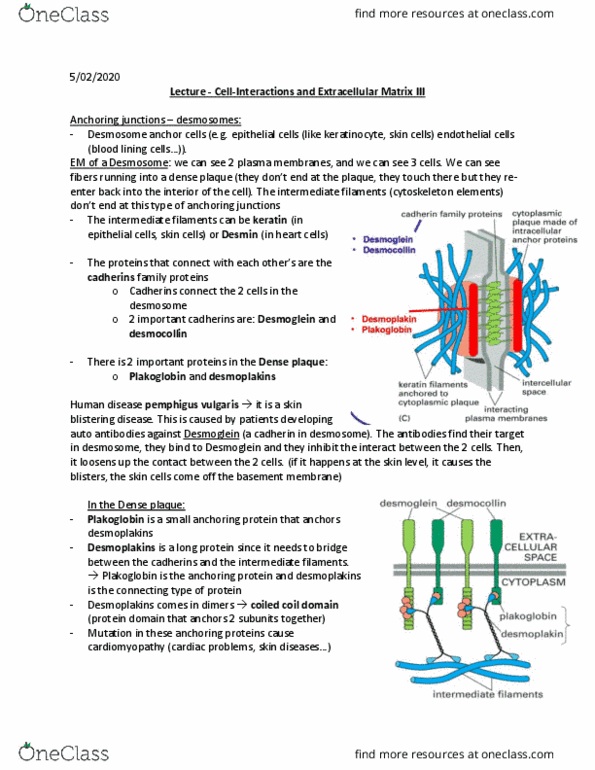
Nerve tissue cadherins have distinct yet overlapping expression patterns. Cadherins: found in the brain of a mouse are the classical cadherins. Cadherin-6: are thought to have a role in synapse formation and stabilization of nerve cells. Proto-cadherins in the brain: they are like antibodies, they can create a variety of cadherins because. Variable region exons: encode for the extracellular region of one type of transmembrane proto-cadherin protein, many promoters associated with each variable exon that are used to. They are identical in their c-terminal regions generate many transcripts. Constant region exons: encoded by separate exons. They are arranged on a proto-cadherin gene cluster. Variability is a result of a combination of differential promoter regions and alternative rna splicing: this is very different in comparison to the antibody diversification, which is done by site-specific recombination. Cytoplasmic tail is is encoded by the three constant-region exons c1, c2, and c3.