MICRB316 Lecture Notes - Lecture 9: Hpk, Dephosphorylation, Signal Transduction

8 Mar 2014
School
Department
Course
Professor
Document Summary
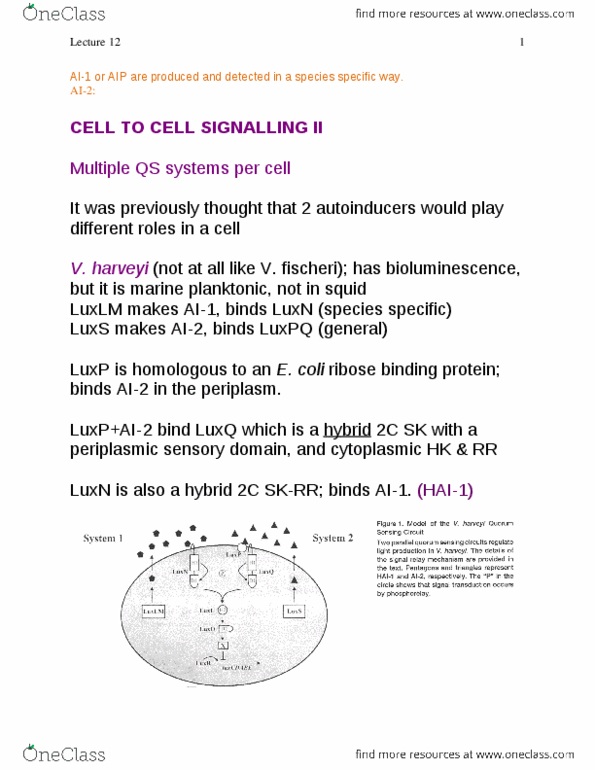
To survive, they must respond rapidly, through: adaption. Therefore, bacteria have evolved mechanisms to sense external conditions & transfer this information to cellular sites where an appropriate response is exerted. This signal transduction generally involves the transfer of phosphoryl groups between proteins. Many of these signals are sensed and transmitted to the tn apparatus by pairs of proteins, called called a 2 component system. These transfer environmental stimuli to bacteria via cognate pairs of. Rr > altered confromation > dna binding by (1) rr itself or (2) other proteins > altered tn. Ntrb is an example of a bi-functional kinase/phosphates. (needs to by able to chut off signal when not needed. Either sk is bi-functional or there is another phophotase involved. Various regulatory mechanisms (some of which act through hpk / sk) regulate intracellular concentration of. When the phosphorylated form of rr (rr~p) reaches a critical level, the dna is bound, leading to rnap recruitment (activation) or interference (repression).