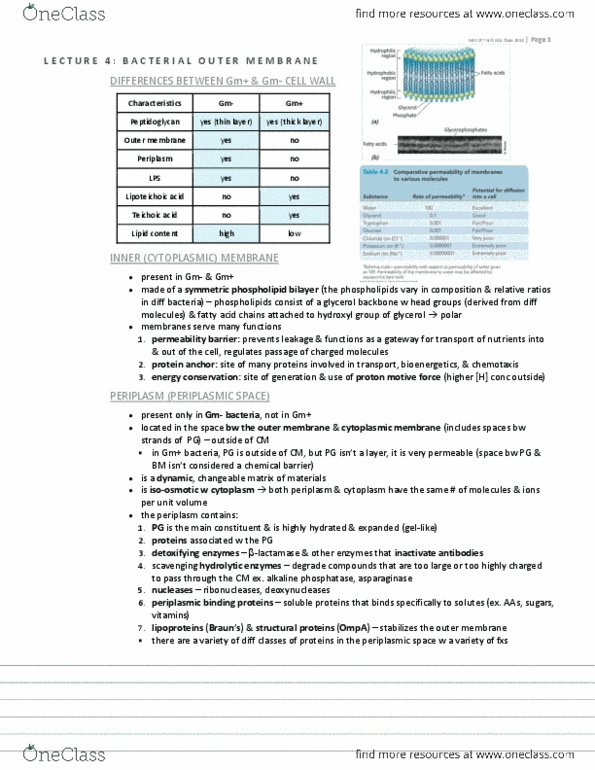
QuestionGroup: #51 Innate immune 1
Assigned as
Homework
51
What is the name of a chemical substance responsible for stimulating an immune response?
A
Pathogen
B
Rotomers
C
Antigen
D
Antibody
E
Troll receptor
Unanswered
Submit
QuestionGroup: #52 Innate immune2
Assigned as
Homework
52
What is the purpose of secondary metabolites produced by normal flora in your body?
A
To aid T cells in fighting local infection
B
To kill infected B cells
C
To help balance electrolytes
D
To inhibit adhesion, and growth of the certain microbes
Unanswered
Submit
QuestionGroup: #53 Innate immune3
Assigned as
Homework
53
The lysozyme removes most bacteria in our foods through the saliva in our mouth, how does it do this?
A
The lysozyme attacks the bacteria through the blood.
B
The lysozyme are not strong enough to get rid of bacteria.
C
The lysozyme breaks down the polysaccharide walls of different bacteria.
D
The lysozyme will tell your body to get rid of the bacteria by vomiting.
Unanswered
Submit
QuestionGroup: #54 Adapt immune 1
Assigned as
Homework
54
In adaptive immunology, the key component is which of the following?
A
Antigen trigger the the production of vaccines.
B
Antibodies will trigger responses in the antigen which will cause an inflamation.
C
Antigens trigger responses of the antibodies which will recognize the antigen.
D
Antibodies will be able to recognize other antibodies and destroy them.
Unanswered
Submit
QuestionGroup: #55 Adapt immune2
Assigned as
Homework
55
An autoimmune induced disorder is...?
A
When the body's immune system attacks a tissue or organ in the body
B
When the immune system is not present
C
When the body attacks the antigens and antibodies of the immune system
D
When the autonomic nervous system attacks the antigen and antibodies of the immune system
Unanswered
Submit
QuestionGroup: #56 Adapt immune3
Assigned as
Homework
56
Antibodies are also called immunoglobulins and constitute part of the blood proteins. How do antibodies work?
A
Agglutination-- they just clump with each other (blocking antigen)
B
Precipitation-- force insolubility and settled out of solution along with antigens
C
Complement fixation-- proteins attach to antigen surface (causing cell lysis)
D
Neutralization-- bind to specific sites to prevent antigen attachment
E
All of the above
Unanswered
Submit
QuestionGroup: #57 Infectious disease
Assigned as
Homework
57
The most deadly STD is HIV, but many other viruses can be deadly. Name the most common sexually transmitted infection.
A
Genital human parisis virus (GHPV)
B
Herpes Simplex
C
Human papilloma virus (HPV)
D
Human pappathe virus tentis (HPVT)
Unanswered
Submit
QuestionGroup: #58 Infectious disease2
Assigned as
Homework
58
Microbes have developed an array of weapons to be able to go from host to host. These virulent factors are divided into several groups, but one important group is the one produced by bacteria. Name the subgroups within this bacterial group.
A
Retretoxin and maitotoxin
B
Retretoxin and butotoxin
C
Endotoxins and exotoxins
D
Lipotoxin and exotoxin
Unanswered
Submit
QuestionGroup: #59 Infectious disease3
Assigned as
Homework
59
Sexually transmitted diseases are very successful at spreading from one person to another. Name one important reason for this fact.
A
Mucous membranes use B cells only and are resistant to antigens
B
Mucous membranes in the sex organs differ from those of the skin in that they allow certain pathogens into the body
C
STDs are transmitted through sex only
D
STDs are usually caused by viruses
Unanswered
Submit
QuestionGroup: #60 HIV/ AIDS
Assigned as
Homework
60
HIV has which of the following characteristics?
A
It has an enzyme called reverse transcriptase which causes the DNA to reverse the nucleotide sequence in messenger RNA.
B
It is a retrovirus meaning that it reverses the order of protein production in that the virus affects the DNA in the cell to make the protein.
C
It is an anti-retrovirus meaning that it attacks the DNA causing the development of new HIV particles.
D
It has a short incubation period which means that the symptoms show up also immediately after infection.