CSB328H1 Lecture Notes - Lecture 10: Bone Morphogenetic Protein 2, Sox2, Growth Factor

25 Jan 2014
School
Department
Course
Professor
Document Summary
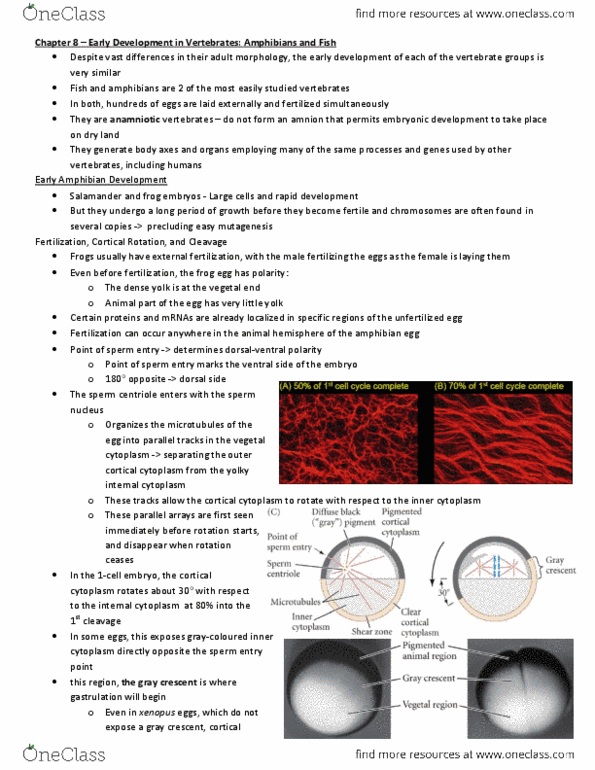
Csb328h1 lisa zhao 2013 | page 1. Fish & amphibians are anamniotic vertebrates (fig 8. 1) Males externally fertilize the eggs as the female lays them. The unfertilized egg has polarity (certain proteins & mrna are already localized) dense yolk at the vegetal pole. Fertilization can occur anywhere in the animal hemisphere. Sperm entry determines the dorsal-ventral polarity sperm entry marks the ventral pole. This exposes the gray-coloured cytoplasm gray crescent region opposite sperm entry point where gastrulation will begin. Amphibian egg undergoes unequal radial holoblastic cleavage (fig 8. 3) Vegetal hemisphere is concentrated w yolk which impedes cleavage. 1st cleavage begins in the animal pole & slowly extends down cortical rotation involves a parallel array of microtubules. 70% of 1st cell cycle complete antibody staining using fluorescent antibodies to tubulin into the vegetal region. While the 1st cleavage furrow is still cleaving the vegetal pole, the 2nd cleavage starts at the animal pole (also meridional)