CSB328H1 Lecture Notes - Lecture 7: Microtubule, Antibody, Subcutaneous Tissue

25 Jan 2014
School
Department
Course
Professor

Document Summary
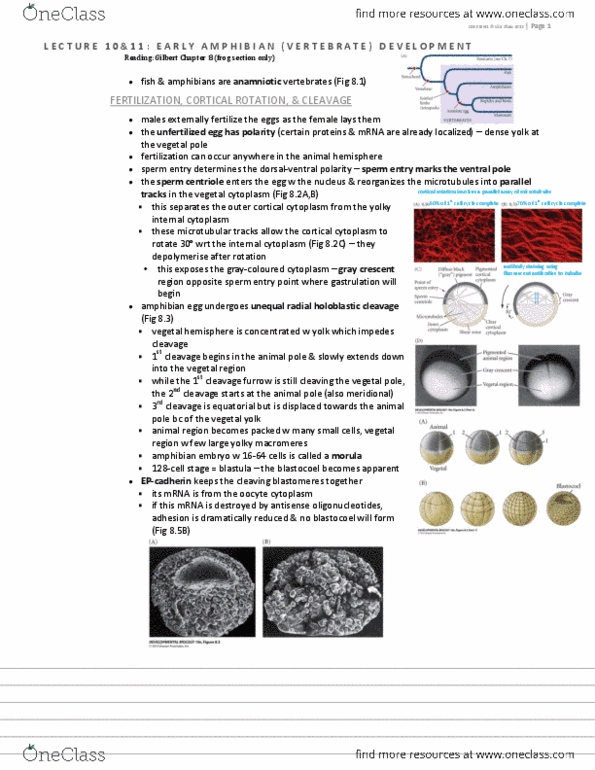
Eukaryotic organism cell contains a nucleus & chromosomes that undergo mitosis. Multicellular eukaryotic organism cells from mitosis remain together & function as a whole. Metazoan animal that undergoes gastrulation (fig 5. 1) Csb328h1 lisa zhao 2013 | page 1. All animals gastrulate & nothing gastrulates that isn"t an animal. 35 metazoan phyla = 35 patterns of animal dvpmt. Cnidarians archeocyte (a somatic cell) can differentiate into all cell types indiv cells can form new organisms, cells are species-specific. Mesoderm no (no organs, circulatory system, nerves, muscles) radial symmetry yes yes no. Page 2 | csb328h1 lisa zhao 2013. Cleavage mitotic divisions, egg cytoplasm divided into smaller nucleated cells. Cleavage-stage cells = blastomeres blastomere: a cell derived from cleavage blastula: embryonic stage composed of blastomeres blastocyst: mammalian blastula blastocoel: cavity in the blastula blastopore: invagination where gastrulation begins. Rate of cell division & placement of cells under control of oocyte (maternal) factors (w exception to mammals)