Biology 2382B Lecture Notes - Lecture 11: Conformational Change, Prometaphase, Actomyosin Ring

24 Aug 2013
School
Department
Course
Professor
Document Summary
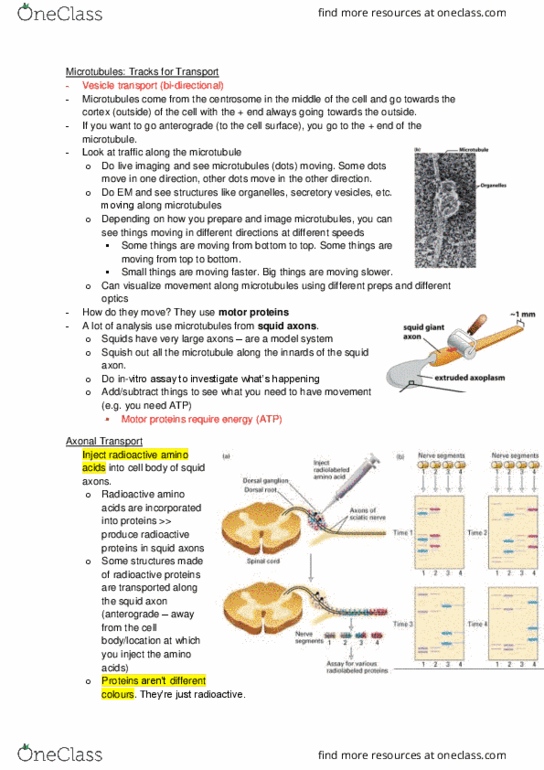
By taking a squid axon (nerve) about 1mm in diameter and squish out all the cytoplasm, we can see all kinds of structures like microtubules. Through adding chemicals like atp, we can observe things moving at different rates and in different directions along microtubules. This is because vesicle transport is bi- directional and the motor proteins doing the transporting require atp energy. The rate of axonal transport in vivo can be determined by radiolabeling and gel electrophoresis. Radioactive amino acids injected into these cell bodies of neurons in experimental animals are incorporated into newly synthesized proteins, which are then transported down the axon to the synapse. These proteins can be identified after gel electrophoresis and autoradiography. The red, blue, and purple dots represent groups of proteins that are transported down the axon at different constant rates, either together or separately. Red moves most rapidly and purple least rapidly.