Hello everyone, i'm working on thesepractices multiple choices for my finals. If you know any of theseanswer, please help. i appreciate your time and your help. Thankyou!!!
1. ___ Which of thefollowing statements about the cytoskeleton is false?
(a) The cytoskeletonis made up of three types of protein filament.
(b) The bacterialcytoskeleton is important for cell division and DNAsegregation.
(c) Protein monomersthat are held together with covalent bonds form cytoskeletalfilaments.
(d) The cytoskeleton of acell can change in response to the environment.
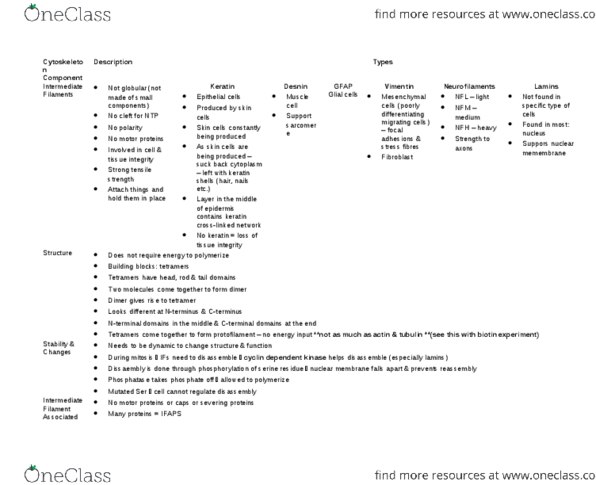
2. ___ Which of thestatements below about intermediate filaments is false?
(a) They can stayintact in cells treated with concentrated salt solutions.
(b) They can befound in the cytoplasm and the nucleus.
(c) They can beanchored to the plasma membrane at cell-cell junction.
(d) Each filament is about10 ?m in diameter.
3. ___ Keratins,neurofilaments, and vimentins are all categories of intermediatefilaments. Which of the following properties below is nottrue of these types of intermediate filaments?
(a) They strengthencells against mechanical stress.
(b) Dimers associateby noncovalent bonding to form a tetramer.
(c) They are foundin the cytoplasm.
(d) Phosphorylation causesdisassembly during every mitotic cycle.
4. ___ Which of thefollowing statements about microtubules is true?
(a) Motor proteinsmove in a directional fashion along microtubules by using theinherent structural polarity of a protofilament.
(b) The centromerenucleates the microtubules of the mitotic spindle.
(c) Becausemicrotubules are subject to dynamic instability, they are used onlyfor transient structures in a cell.
(d) ATP hydrolysis by atubulin heterodimer is important for controlling the growth of amicrotubule.
5. ___ Thehydrolysis of GTP to GDP carried out by tubulin molecules________________.
(a) provides theenergy needed for tubulin to polymerize
(b) occurs becausethe pool of free GDP has run out
(c) tips the balancein favor of microtubule assembly
(d) allows the behavior ofmicrotubules called dynamic instability
6. ___ The microtubulesin a cell form a structural framework that can have all thefollowing functions except which of the following?
(a) holding internalorganelles such as the Golgi apparatus in particular positions inthe cell
(b) creating longthin cytoplasmic extensions that protrude from one side of thecell
(c) strengtheningthe plasma membrane
(d) moving materials fromone place to another inside a cell
7. ___ Which of thefollowing statements about organellar movement in the cell isfalse?
(a) Organellesundergo saltatory movement in the cell.
(b) Only themicrotubule cytoskeleton is involved in organellar movement.
(c) Motor proteinsinvolved in organellar movement use ATP hydrolysis for energy.
(d) Organelles areattached to the tail domain of motor proteins.
8. ___ Which of thefollowing statements is correct?
Kinesins and dyneins____________________.
(a) have tails thatbind to the filaments
(b) move along bothmicrotubules and actin filaments
(c) often move inopposite directions to each other
(d) derive their energyfrom GTP hydrolysis
9. ___ Cell movementinvolves the coordination of many events in the cell. Which of thefollowing phenomena are not required for cell motility?
(a) Myosin-mediatedcontraction at the rear of the moving cell.
(b) Integrinassociation with the extracellular environment.
(c) Nucleation ofnew actin filaments.
(d) Release ofCa2+ from the sarcoplasmic reticulum.
10. ___Which of the followingstructures shorten during muscle contraction?
(a) myosinfilaments
(b) flagella
(c) sarcomeres
(d) actin filaments
7. ___ Referring to Figure Q16-29,which of the following statements below is false?
(a) A constitutivelyactive mutant form of PKA in skeletal muscle cells would lead to adecrease in the amount of unphosphorylated phosphorylasekinase.
(b) A constitutivelyactive mutant form of PKA in skeletal muscle cells would notincrease the affinity of adrenaline for the adrenergicreceptor.
(c) A constitutivelyactive mutant form of PKA in skeletal muscle cells would lead to anexcess in the amount of glucose available.
(d) A constitutivelyactive mutant form of PKA in skeletal muscle cells would lead to anexcess in the amount of glycogen available.
8. ___ Match the targetof the G protein with the appropriate signaling outcome.
adenylyl cyclase________ A. cleavageof inositol phospholipids
ion channels_________ B.increase in cAMP levels
phospholipase C_________ C. changes in membranepotential