BMS 460 Lecture Notes - Intermediate Filament, Cytoskeleton, Rigor Mortis

26 Apr 2014
School
Department
Course
Professor
Document Summary
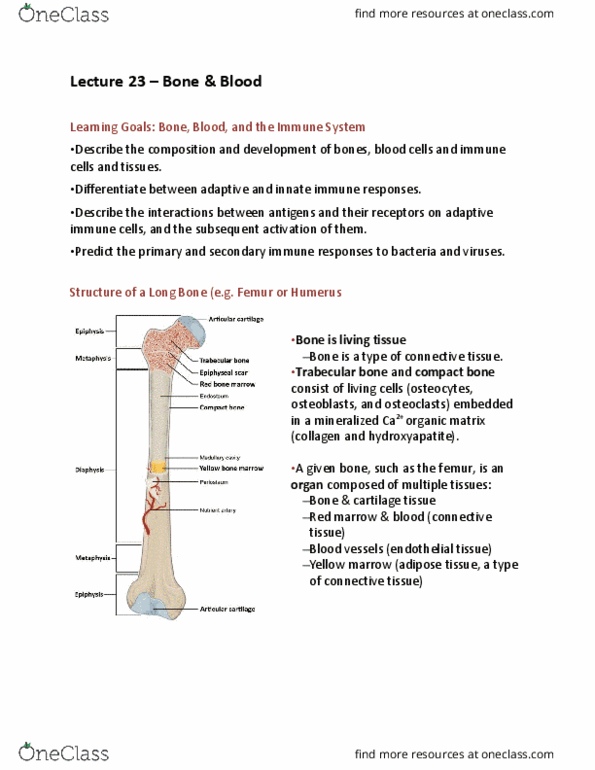
Cartilage and bone are specialized forms of connective tissue derived from embryonic mesenchyme. Both consist of cells embedded in an extracellular matrix. Cartilage matrix is highly hydrated, being 70 75% water. The rest of the matrix is composed of collagen (15 20%), for tensile strength, and proteoglycans (2 10%) for resistance. It is avascular and has no nerve or lymphatic supply. Bone is the calcified component of the skeleton. The matrix of bone consists of collagen embedded in a ground substance on which is deposited a complex inorganic mineral, hydroxyapatite. Compared with cartilage, bone has a higher metabolic rate, is richly vascularized, and receives up to 10% of cardiac output. Bone has good regenerative potential for self-repair throughout life, whereas cartilage has a very limited capacity for regeneration in response to traumatic injury or disease. Bone develops with or without a cartilage intermediate.