BIOL 4004 Lecture Notes - Lecture 1: Threonine, Digestion, Proteoglycan

Document Summary
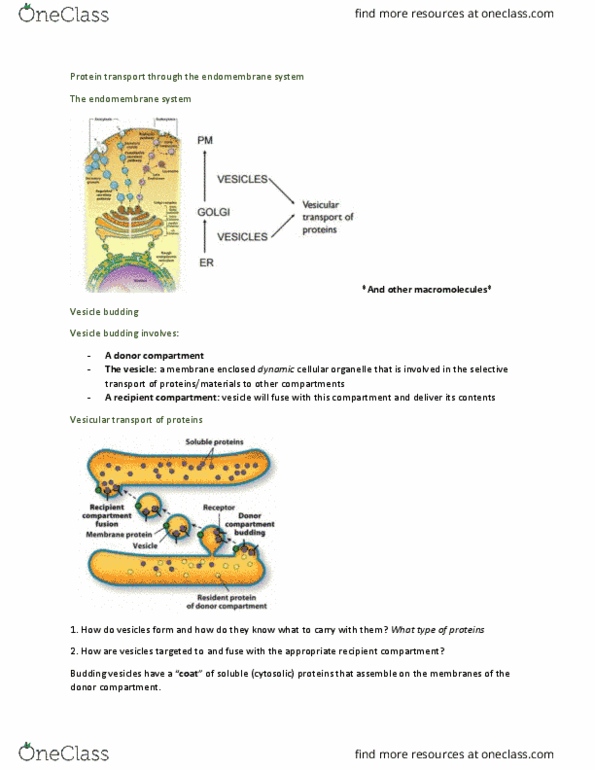
Mbc5 study guide chapter 13 (intracellular vesicular traffic) Thus far, we have considered protein-targeting mechanisms that involve receptor binding and the subsequent movement of proteins through channels, either posttranslationally or cotranslationally. This chapter considers another way that proteins move from compartment to compartment via membrane vesicles. It occurs between various cellular compartments as depicted in figure 13-3. The molecular mechanisms of membrane transport and the. Figure 13-2 gives a general overview of vesicular transport. A membrane vesicle buds from a donor compartment and later fuses with a target compartment. As shown in figure 13-3, this type of vesicular transport occurs throughout the secretory pathway. The vesicles that bud from donor membranes are termed coated vesicles because they are coated with particular protein molecules. The protein coats promote the budding process and also play a role in the fusion of the vesicle with the correct target membrane. The three types, clathrin, copi , and copii, are shown in figure 13-4.