BIOL 1090 Study Guide - Final Guide: Nuclear Pore, Nuclear Lamina, Nuclear Membrane

12 Apr 2013
School
Department
Course
Professor



Document Summary
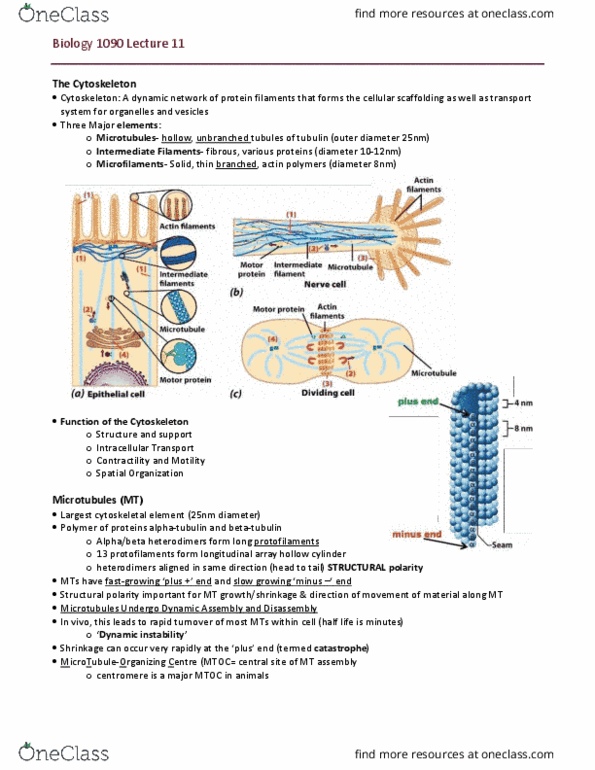
Dynamic network of filaments and tubules that extend through the cytosol. Structural polarity is important for mt growth/shrinkage direction of movement of material along mt: shrinkage can occur very rapidly at the plus" end (catastrophe, dynamic instability. Microtubule-organizing center-mtoc central site of mt assembly) Non-motor: control mt organization in cytosol. Motor: two types (can power intracellular transport): Kinesin (plus end directed) and dynein (minus end directed: use atp to generate force, can move material along mt track", can generate sliding force between mts. Contains a-helical domains: wrap around each other forming a coiled dimer. Monomers are aligned in parallel if dimers are polar. Assembly: g-actin polymerizes reversibly, nucleation (slow): Polymerization/depolymerization and structure/organization of f-actin filaments are regulated by actin-binding proteins: filaments can be lose or tight. Myosin is a motor protein associated with it: large family of proteins, most move its toward + end, divide into 2 groups: Generate force and contribute to mobility in non-muscle cells.