BIOCHEM 2B03 Chapter Notes - Chapter 30: Ribonucleoprotein, Hydrolase, Hydrophobe

6 Sep 2013
School
Department
Course
Professor
Document Summary
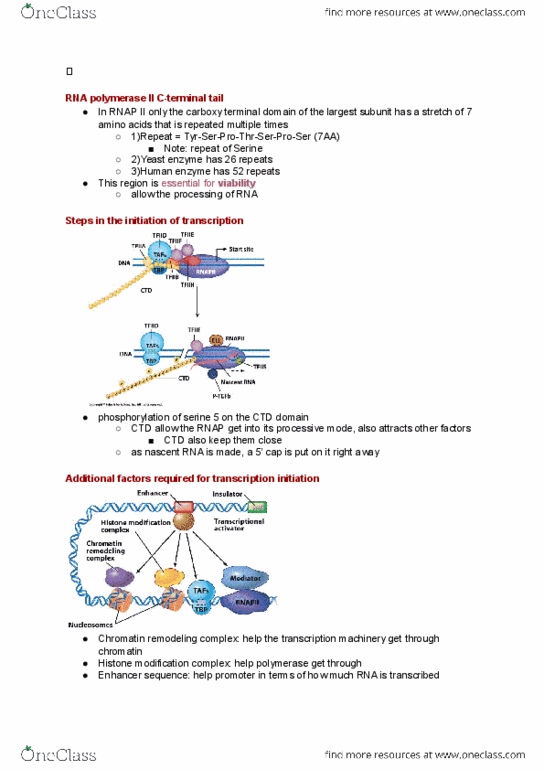
Ctd of rpb1 is essential to the function of rnap ii. Rnap ii only initiates transcription when the ctd is not phosphorylated. Elongation proceeds only after phosphorylation of ctd, suggesting phosphorylation triggers initiation to elongation conversion. Following termination, a phosphatase recycles rnap ii for initiation. Ctd also play a prominent role in orchestrating subsequent events for processing the transcript (capping, splicing, General structure of amino acids: c is a chiral center (with one exception) In 1960 you couldn"t sequence dna but could sequence protein. Rna has four bases c, g, a, u. 42 ways of combining these in pairs. Three bases codes for one amino acid. Base sequence is read from a fixed starting point without punctuation. Code is degenerate (each amino acid can be coded for by several triplets) Prime them with synthetic rnas of defined sequence. Note: termination codons, start codon, redundancy: trp has the only unique codon. Codons triplets of bases in mrna; read 5" 3".