BIO120H1 Chapter 15: Bio Chapter 15.doc



36
BIO120H1 Full Course Notes
Verified Note
36 documents
Document Summary
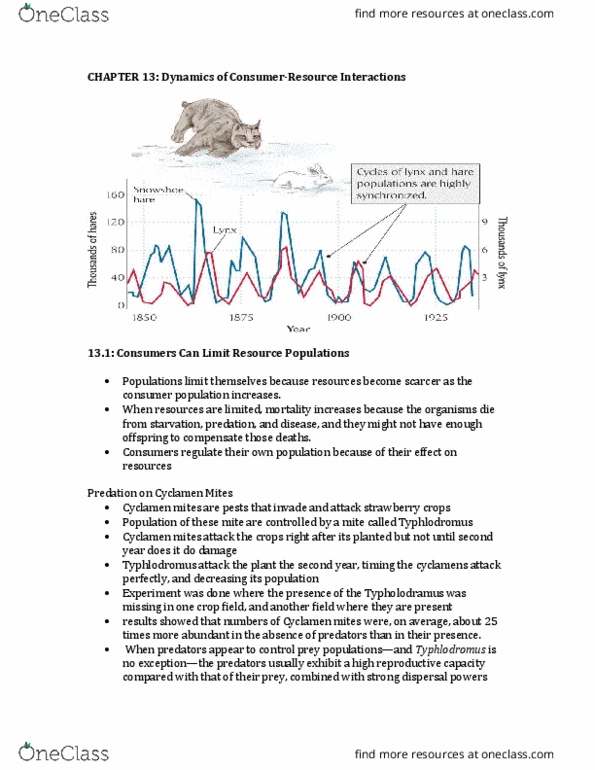
Chapter 15 dynamics of consumer resource interactions (302-311, b; 312-315, a; 315-320, c; 321-322, a/b) Consumers can exert a substantial influence on resource populations. Consider the example of cyclamen mites (pest to strawberry plants) and their natural predators (also mites). By experimentation, it was found that cyclamen were 25 times more abundant in the absence of predator mites. Generally the degree to which a resource population may be controlled by consumers depends on their respective life histories. This influence in not exclusive to animals, in many instances, invasive plant species have been brought under control by introducing consumer species. These interactions can be modeled with relatively stable cycles (1: high prey, low predator, 2: high predator, low prey). The periods of these cycles depend on the reproductive cycles of specific species (this introduces time delays) and the environment (assuming it follows periodic cycles of favourable vs. unfavourable conditions, predator-prey cycles are still quite predictable).