BIO130H1 Chapter Notes -Guanine, Tata Box, Cleavage And Polyadenylation Specificity Factor



31
BIO130H1 Full Course Notes
Verified Note
31 documents
Document Summary
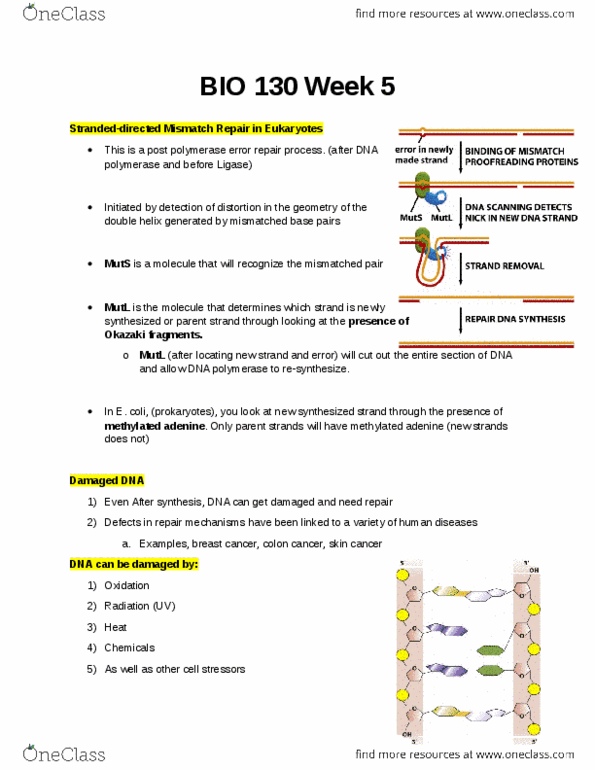
A strand-directed mismatch repair system removes replication errors that escape from the replication. Machine: the strand-directed mismatch repair system detects the potential for distortion in the. Dna helix from the misfit between noncomplementary base pairs. In order to be effective, a correction mechanism must be able to distinguish and remove the mismatched nucleotide only on the newly synthesized strand, where the replication error occurred. Dna polymerases, using these extensions as a template to synthesize the complementary strand. Telomere length is regulated by cells and organisms. Coupling dna repair to transcription ensures that the cell"s most important dna is efficiently. Dna: dna is repaired using the sister chromatid as a template. How cells read the genome: from dna to protein. It seems like these two polymerase enzymes have arisen independently. Signals encoded in dna tell rna polymerase where to start and stop. If a protein is used in abundance, its promoter sequence is much stronger than a protein used rarely.