PHGY 209 Lecture Notes - Lecture 7: Basilar Membrane, Oval Window, Tectorial Membrane

Document Summary
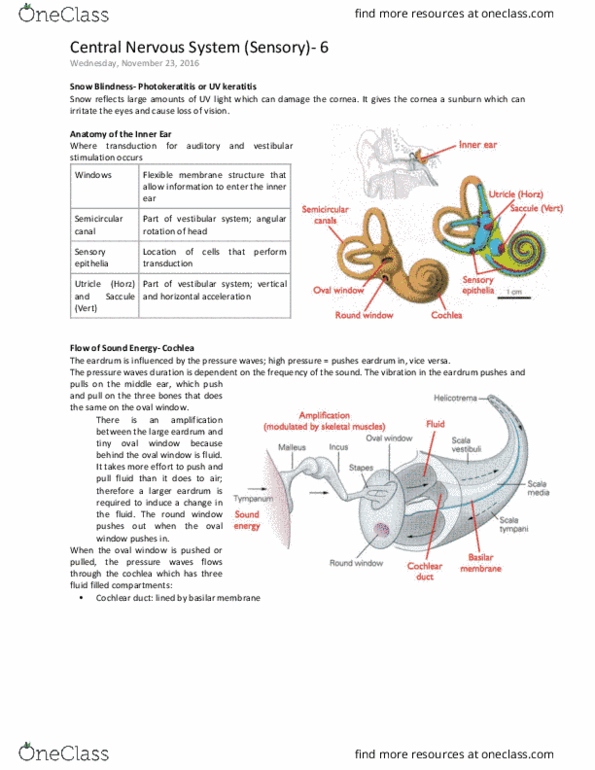
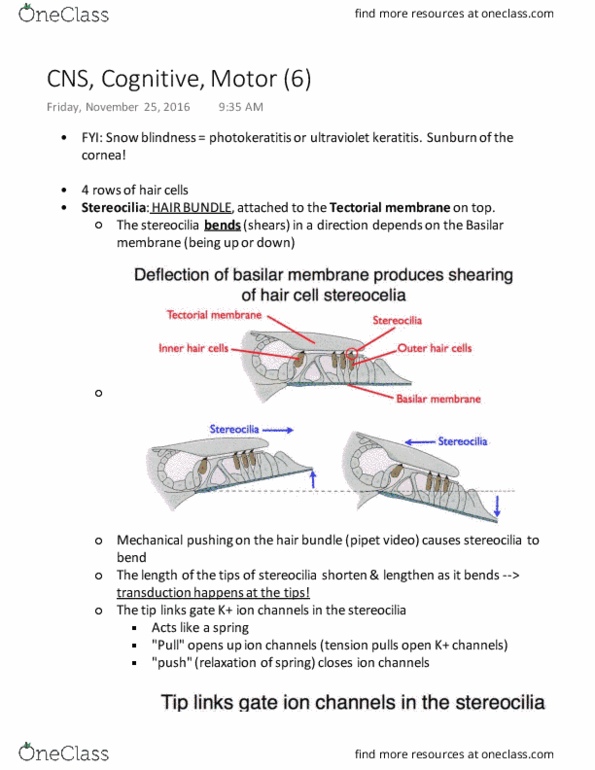
It turns out that the basilar membrane is critical for the transduction process. The basilar membrane is stiff near the oval window and as we get farther away from it, the membrane gets more flexible: low frequencies vibrate towards the end of the basilar membrane, away from the oval window. High frequencies, on the other hand, vibrate closer to the oval window. If we have a mixture of frequencies, we have vibrations at different places: what the basilar membrane is doing is extracting the frequency of the sound. In a 1khz sound, the time these pressure waves flow by your head is every 1 milisecond: remember this is the cochlea which we"ve unrolled. Video of a computer model of the basilar membrane. The model was made by a researcher named dr. james hudspeth, from. Over the last 20 years, he has been one of the leaders at revealing the transduction process of the auditory system.