NROC69H3 Lecture Notes - Lecture 2: Vesicle Fusion, Porosome, Chemical Synapse

Document Summary
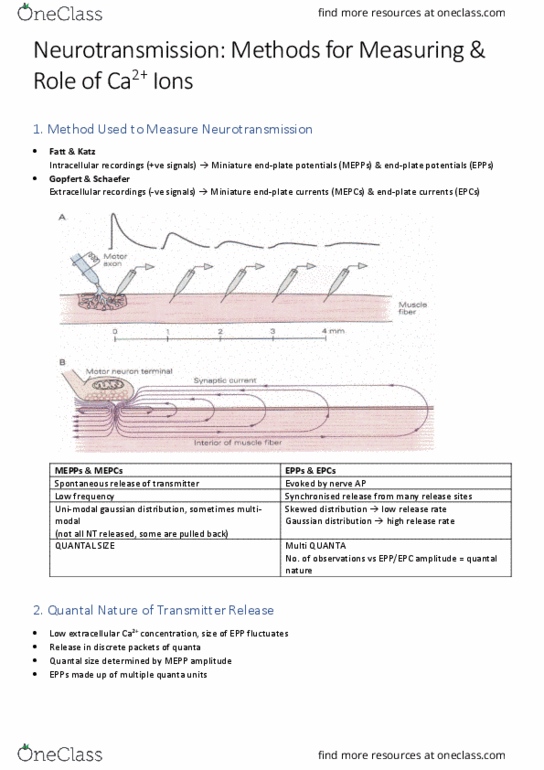
Action potential down an axon to the nerve terminal. Opening of voltage-gated ca 2+ channels in the presynaptic terminal. This allows the influx of ca 2+ Neurotransmitters bind to plasma membrane and release them into synaptic cleft. Post synaptic receptor neurotransmitters gets diffused ** happens in less than 1 ms. Other neurotransmitters either get taken back up or they get degraded. Some of them move away in other areas ** volume transmission. They then diffuse across to bind to postsynaptic receptors either ionotropic or metabotropic. Fatt & katz: discovered a spontaneous miniature end plate potential (mepps) of ~0. 5 mv. They found this spontaneous mepp at the neuromuscular junction of a frog during resting state. There was no stimulation but a 0. 5 mv at resting phase. Presynaptic depolarization stimulation of the motor neuron increased the frequency occurrence of mepp but it did not change their amplitude suggesting 1 ach binding of 1 ach receptor.