PSL201Y1 Lecture Notes - Extracellular Fluid, Carbohydrate, Endosome

Document Summary
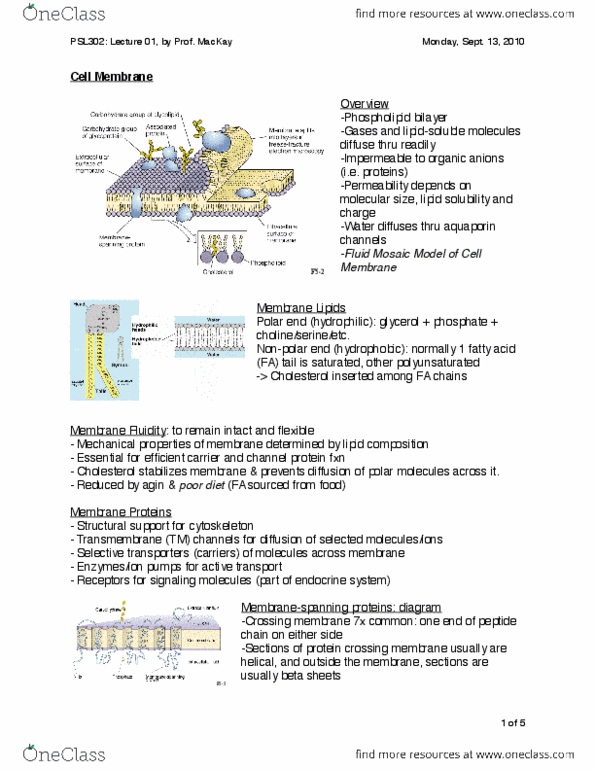
Charge: membrane = permeable to a substance if the substance can cross by any means, permeable. So diffuse thru readily (won"t let thru) to: Need prot channels or carriers to cross. Structure fluid-mosaic model: phospholipid b ilayer. Polar head (inner+outer surface): glycerol phosphate (hydrophilic) choline/serine or other amino acids. Splits into layers in freeze fracture electron microscopy. Important for cell identification (ex. immune responses), cell adhesion. Membrane fluidity: determined by lipid composition, essential for working protein carriers and channels (allowing shape changes for function, reduced by aging and poor diet. Wrong proportion of amino+fatty acids in membrane: c holesterol. Prevents diffusion of polar molecules across it. Membrane proteins (embedded) for active transport for signaling molec in metabolic processes structural support for anchoring to cytoskeleton in cell interior stabilization: receptors, enzymes/ion pumps, membrane-spanning proteins. Cross membrane multiple times (most commonly 6 or 7) Loops on outer surfaces often attached to carb chains. Nh2 at one end, cooh at the other (glycocalyx)