Biology 1001A Lecture Notes - Okazaki Fragments, Bromine, Release Factor

10 Apr 2013
School
Department
Course
Professor
Document Summary
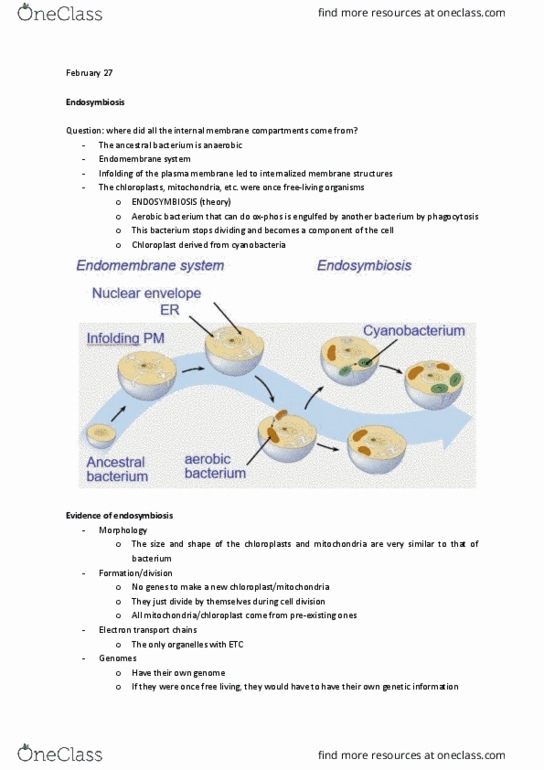
Endomembrane system is a collection of inter-related internal membranous sacs that divide into functional and structural compartments called organelles. Major membrane components are: the nuclear envelope, the er, the golgi complex. Infolding of the plasma membrane is believed to be responsible for the evolution of all these structures. In the cell lines leading from prokaryotes to eukaryotes, pockets of the plasma membrane may have extended inward and surrounded the nuclear region origin of mitochondria and chloroplasts: Prokaryotic ancestors of modern mitochondria and chloroplasts were engulfed by larger prokaryotic cells, forming a mutually advantageous relationship called a symbiosis. And over time the host cell and the endosymbionts became inseparable parts of the same organism. Descendants of ancient free-living prokaryotes evidence supporting theory of endosymbiosis. Morphology: the form or shape of both mitochondria and chloroplasts is similar to that of a prokaryotic cell.