CAS BI 315 Lecture Notes - Lecture 63: Spliceosome, Ceramide, Eif2B


- snoRNPs and snRNPs have
different protein and RNAs.
- In the nucleolus, ribosomal
proteins enter and the pre-rRNA is
spliced (BUT 5s rRNA comes from
elsewhere in the nucleus) and
partly assembled into the 60S
subunit and 40S subunit.
Remember! The subunits come
together in the cytosol during
translation.
-Cajal bodies in the nucleus. Cajal
bodies are involved in assembly of snRNPs and other RNA-protein complexes.
- Cajal bodies appear to play a role in assembly of telomerase, which replicates the ends
of chromosomal DNA. We know this using immunocytochemistry - Using specific
antibodies against specific proteins (i.e. telomerase or proteins in the spliceosome
complex).
- Nuclear speckles also contain splicing factors. Cajal Bodies are where the spliceosomes
are assembled and speckles are where splicing takes place.
Protein Sorting and Transport
- How do cells traffic (or sort) proteins to their destined locations (or compartments)?
- Remember! Translation always starts on free ribosomes and then gets transported to the
RER
- Nuclear Proteins are imported into the Nucleus by importins following translation on “free
ribosomes”. Proteins destined for many compartments are imported following translation
on free ribosomes. Important: If the protein is destined to the Nucleus, Peroxisomes,
Mitochondria, or Chloroplasts, then DO NOT go to the ER. Always on free ribosomes
- BUT, Proteins destined for several other compartments are trafficked differently.
Examples: plasma membrane proteins, secreted proteins, ER proteins, Golgi proteins,
Lysosomal proteins. These all have a common pathway.
- How are these protein trafficked (or sorted) to their respective compartments? Via the
secretory Pathway
- Key Study: Examined the path through which newly synthesized proteins are secreted
from cells from pancreatic cells.
1. “Pulse-labeled” newly synthesized proteins with radioactive “hot” amino acids.
They looked at this afterwards with a film.
2. The radioactive AAs solution would be removed and then “chased” with
nonradioactive “cold” amino acids.
3. Tracked the location of the radioactive protein through time.
- Pancreatic cells are polarized and they look like little beakers. The RER is at the
bottom and the Golgi is in the middle and secretory vesicles are up top. New
proteins that are made are labelled since they have radioactive AAs. Translation
happens within the first 3 minutes (3-min label) and the proteins are in the RER.

In 7 minutes (7-min chase) the proteins are no longer in the RER but rather in the
Golgi. So the proteins also move relatively fast to Golgi. Proteins spend a long
time in the Golgi though. 120 minutes later (120-min chase) the proteins can be
seen in the secretory vesicles and secreted out of the cells.
- The Secretory Pathway: ER → Golgi → secretory vesicles → cell exterior
- First Step: ER
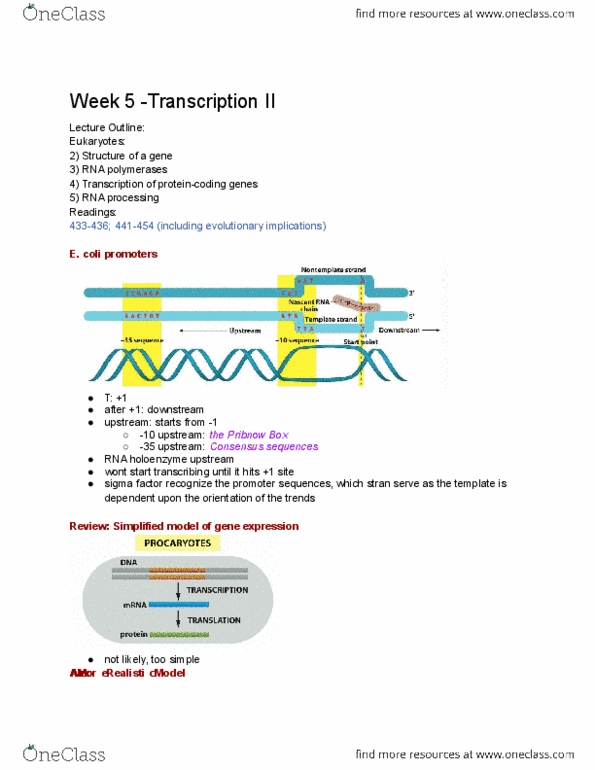
- How does translation get to the ER? The Signal
Hypothesis: Remember! 1) Translation of secreted
proteins takes place on ER-bound ribosomes 2)
the protein was transferred across the membrane
during synthesis 3) Secreted protein had 20 AAs
missing at its N-terminus compared to protein
translated in vitro
on “free ribosomes”.
- AA sequence at the N-terminus of a growing
polypeptide chain targets the ribosome to the ER
→ “Signal Sequence”
-First experimental approach to test the “Signal
Hypothesis”:
- The got microsomes via cell disruption
(microsomes are basically little balls of SER and RER). They centrifuged
the microsomes to get the heavier RERs. Then they did protein
translation using RER and free ribosomes. They found that the free
ribosomes made longer IgG proteins (20-25 AAs longer) than the RER
microsomes.
- Signal sequences are recognized by SRP (signal recognition particle)(SRPs
aren’t proteins. They’re part rRNA and part protein), which directs “nascent”
proteins (proteins in the process of being translated by a ribosome) to the ER via
interactions with the SRP receptor.
- SRP identifies and binds to signal sequence → SRP takes the whole ribosome
and protein complex to the RER and binds to a SRP receptor (part of Protein
Translocation complex) → SRP goes away and protein begins synthesizing into
the RER → Translocon (translocates the protein into the ER) has a little flap that
prevents protein from going into the ER until signal peptidase cuts off the signal
sequence. Insertion of the signal sequence opens the translocon by moving a
plug away from the channel. Elongation of the polypeptide drives its transfer
through the translocon. Ie LDL receptor has ~25 AAs signal sequence.
-Signal sequences (about 20 AAs) include a stretch of hydrophobic residues,
(Remember! NLS is mostly basic AAs) and are usually located at the amino
terminus of the polypeptide chain.
- N-terminus signal sequences target proteins for “co-translational” (starts in
cytosol and ends in ER) import into the ER
Document Summary
Cajal bodies are where the spliceosomes are assembled and speckles are where splicing takes place. Protein sorting and transport snornps and snrnps have different protein and rnas. In the nucleolus, ribosomal proteins enter and the pre-rrna is spliced (but 5s rrna comes from elsewhere in the nucleus) and partly assembled into the 60s subunit and 40s subunit. The subunits come together in the cytosol during translation . bodies are involved in assembly of snrnps and other rna-protein complexes. Cajal bodies appear to play a role in assembly of telomerase, which replicates the ends of chromosomal dna. We know this using immunocytochemistry - using specific antibodies against specific proteins (i. e. telomerase or proteins in the spliceosome complex). Translation always starts on free ribosomes and then gets transported to the. Nuclear proteins are imported into the nucleus by importins following translation on free ribosomes . Proteins destined for many compartments are imported following translation on free ribosomes.