BCH210H1 Study Guide - Final Guide: Lipoprotein, Phosphoglucomutase, Triglyceride

13 Jul 2013
School
Department
Course
Professor


49
BCH210H1 Full Course Notes
Verified Note
49 documents
Document Summary
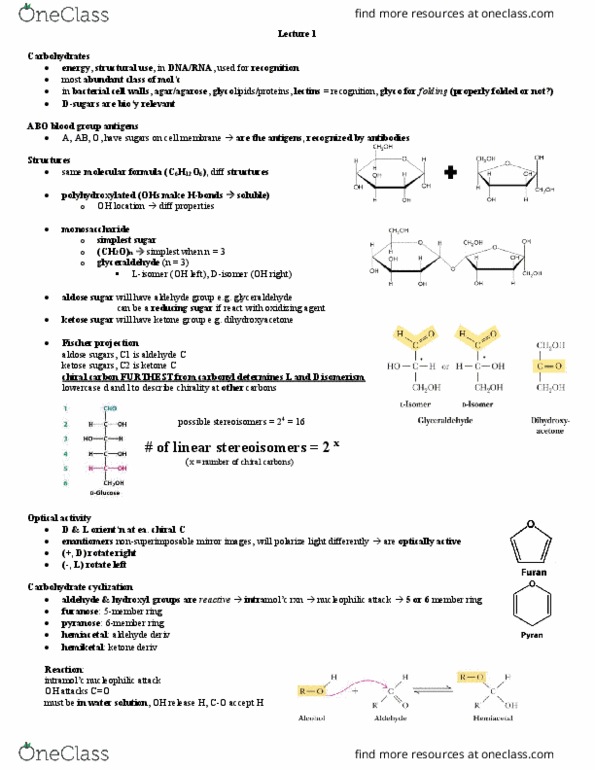
D-glyceraldehyde is a 3c sugar with an aldehyde and a ch2oh on the end. Ketoses are numbered relative to the position of the ketose. The aldehyde and ketone carbonyl undergo nucleophilic attack by the oh group to form pyran or furan rings. Hemiacetal: the reaction of an aldehyde with hor" leads to the. Mutarotation: attack of the carbonyl by oh in the structure. Alpha: the new oh group is on the opposite side as the ch2oh. Oh is axial and below the plane of the ring. The oh is equatorial and above the plane of the ring. Anomer: identical except for the position of the oh at the new chiral centre. In order for something to be a reducing sugar, the free hydroxyl must be adjacent to the o molecule within the ring. Condensation reaction: releasing h2o to form a glycosidic bond. Cellubiose = glucose x 2, but with a b-1, 4 linkage.