CSB331H1 Lecture Notes - Lecture 7: Fibrin, Phosphoinositide 3-Kinase, Atp Hydrolysis

8 Jun 2018
School
Department
Course
Professor


Lecture 7: Actin Dynamics Continued
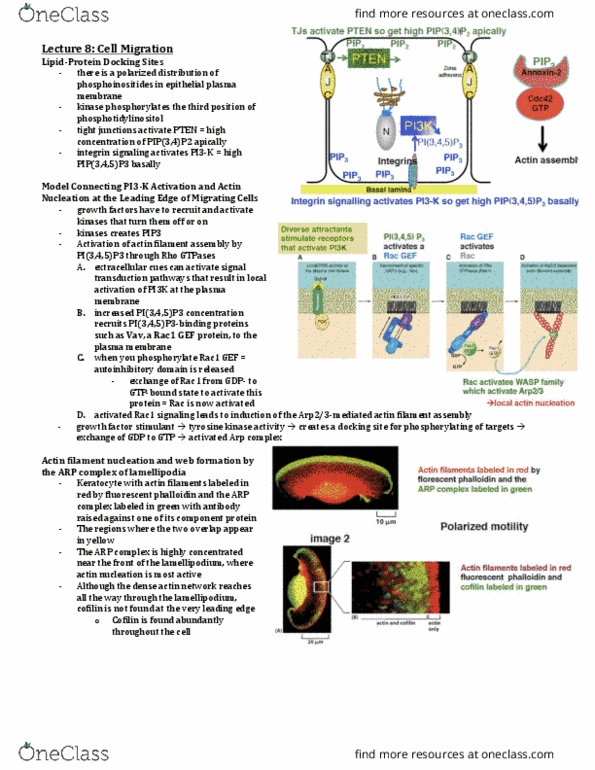
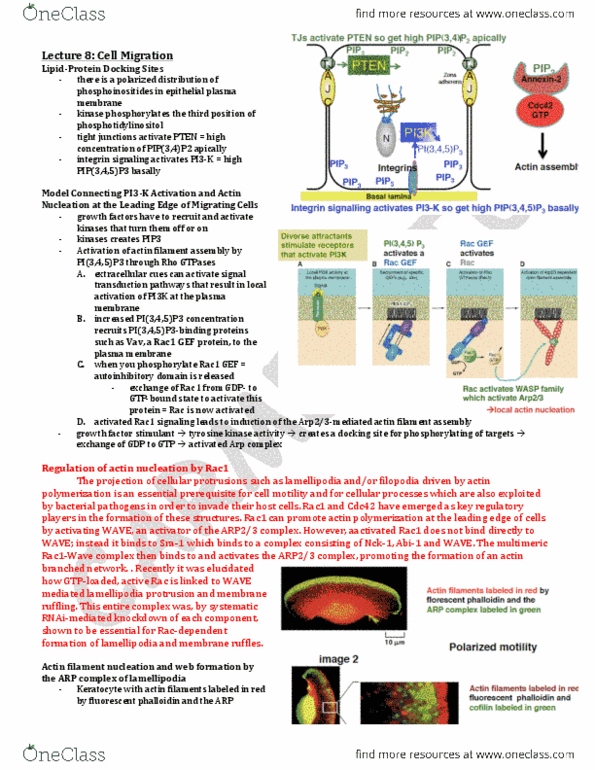
A model for protrusion of the actin meshwork at the leading edge
Arp 2/3 complex mimics the barbed end
o Arp Complex: nucleates actin filament growth from the
minus end, allowing rapid elongation at the plus end
o Can also attach to the side of another actin filament
while remaining bound to the minus end of a filament
that it has nucleated, and can make a treelike web.
o Arp complex is associated with structures at the leading
edge of migrating cells – the complex is localized in
regions of rapid actin filament growth such as
lamellipodia, and intracellular signaling molecules and
components of cytosolic face of the plasma membrane
regulate its nucleating activity
o Conserved complex is also involved in actin filament
nucleation near the plasma membrane – required to
form cortical actin patches
Actin Treadmilling – polymerization at the leading edge and depolymerization at the trailing end
o Moves the actin away from its original site
o Ex: sheets of epithelial moving; like wound repair, keratinocytes would move to reseal the wound
Regulation of nucleation event with an activator (N-WASP) which itself needs to be activated by Rho factors which then need to be
activated by things like growth signaling
Actin is a powerful molecule and therefore must be tightly regulated by layers of regulation
o These proteins are multi-functional; not practical to name proteins
after initial discovery because it limits perspective of their function
Filaments are organized into higher-order structures:
Two types of actin filament arrays:
1. Bundles
2. Web/Gel like networks
Accordingly there are two classes of actin cross-linking proteins – these are under regulation
and have multiple functions
1. Bundling proteins – alpha-actinin, fimbrin (villin, fascin)
2. Gel-forming proteins – filamin, spectrin
Modular Structures of Four Actin-Cross-Linking-Proteins
Each protein has two actin-binding sites (red) and are related in sequence
Gel-forming:
Filamin has two actin-binding sites with a V-shaped linkage between them - cross links actin filaments oriented at right angles
o Can bind to activating scaffolds that connect it to integrins – important role in formation of focal contact points
o One of its roles is to stabilize the gel-like network – have interactions with adaptor molecules or kinases that interact with
the plasma membrane
o Get a separation between he dimer at the binding end and more covalent association at the other end
Stress fibers are contractile and exert tension.
Filopodia are spike-like projections of the
plasma membrane that allow a cell move.. The
cortext underlies the plasma membrane.

o If you want to form a gel, you don’t want tight associations – able to splice them apart to interact with distant filaments
o Cells require the actin gel to formed by filamin in order to extend the thin sheet-like membrane projections called
lamellipodia that can help them crawl across solid surfaces
Spectrin is a long, flexible protein - tetramer that has two alpha and two beta subunits – two actin bindings sites
o In RBC, concentrated just beneath the plasma membrane where it forms a 2D web held together by short actin filaments
o Spectrin links this web to the plasma membrane because it has separate binding sites for peripheral membrane proteins
o Creates a stiff cell cortex that provides mechanical support for the overlying plasma membrane, and allow RBC to spring
back to original shape after squeezing through capillaries
Bundling:
Fimbrin has two directly adjacent actin-binding sites – closely packed and
aligned with the same polarity
o Crosslinks actin filaments into tight bundles which exclude myosin
o So not contractile (because so tight)
The two actin binding sites in alpha-actinin are separated by a spacer
o Homodimer, cross links actin filaments into loose bundles, which
allow the motor protein myosin II to participate in the assembly
o (Myosin is a motor protein – Myosin II is responsible for muscle
contraction)
o Polarity in opposite directions, anti-parallel so myosin motors can
move filaments
Since different packing, they discourage each other, so are mutually exclusive
Note: motors are actin binding, so if they are too locked in (too tight) they would be no movement – needs to be interaction that
promotes bundling that does not restrict the sliding of the filaments past each other
o Not locked in otherwise, would not have any contractile activity but it does help organize the filament so the myosin
motor retains its activity in a maximum way
Microvilli – bundles of parallel actin filaments cross-linked by fimbrin (and villin)
A microvillus contains fimbrin and villin – depends on specific tissues
A bundle of parallel microvillanet s cross-linked by actin-bundling proteins make up the core
Lateral sidewarms (composed of myosin I and the Calcium binding protein calmodulin) connect
the sides of the actin filament bundles to the overlying plasma membrane
All plus ends of the actin filaments are at the tip of the microvillus, where they are embedded in
an amorphous, densely staining substance of unknown composition
The tail domain is where myosin motors give functional diversity – where you vary between
family members of myosin motors
o Why would you have myosin motors tethered to the plasma membrane?
o They are in dynamic flux and promote movement along the microfilaments and
transporting stuff
Freeze-fracture electron micrograph of the apical surface of an intestinal epithelial cell
B) Actin bundles from the microvilli extend down into the cell and are rooted in
the terminal web, where they are linked together by a complex set of proteins
that includes spectrin and myosin II. Below the terminal web is a layer of
intermediate filaments.
o Dense amorphous region contains memebers of formin family and its
activator (packaged together)
o At surface of embryo, would see cells with cilia coming from the
surface surrounded by non-cilliated cells
o Cilia beat rhythmically and in the same direction (planar polarity) and
pass oxygen on the surface
o Cilia and microvilli are very dynamic and constantly disassemble and
turn over (not stable)
Document Summary
A model for protrusion of the actin meshwork at the leading edge. Arp complex: nucleates actin filament growth from the minus end, allowing rapid elongation at the plus end. Can also attach to the side of another actin filament while remaining bound to the minus end of a filament that it has nucleated, and can make a treelike web. Conserved complex is also involved in actin filament nucleation near the plasma membrane required to form cortical actin patches. Actin treadmilling polymerization at the leading edge and depolymerization at the trailing end: moves the actin away from its original site. Ex: sheets of epithelial moving; like wound repair, keratinocytes would move to reseal the wound. Regulation of nucleation event with an activator (n-wasp) which itself needs to be activated by rho factors which then need to be activated by things like growth signaling. Actin is a powerful molecule and therefore must be tightly regulated by layers of regulation.