PSL300H1 Lecture Notes - Lecture 11: Parietal Lobe, Cytochrome C Oxidase, Color Vision

Document Summary
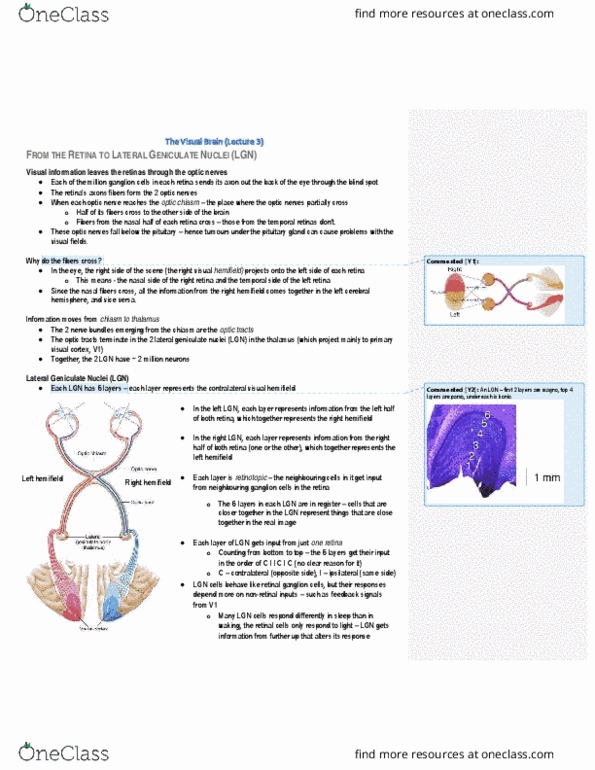
Optic chiasm: each retina sees" both visual hemifields, but each half of brain processes only contralateral field. Sc/tectum in midbrain), and accessory optic system for motion detection (a lot of m stream input) And all info of left hemifield in left eye must. Joining together on one side all info of. Like in s1 w magnification of areas of fingers. Point is dividing up layers btwn the 2 eyes. If take radial axis thru all layers, the neurons in each layer along it will respond to same spot in contrlateral visual field. Right lgn mapping left half of visual world all nerons along radial axis respond to same place in visual field. Organization - retinotopic mapping, each neuron has place in layer, lined up across the layers. Fovea, central 3 degrees (actually 1. 5 since hemifield) of physical visual field. Blown up disproportionately in v1 over a third of v1 devoted to this central portion of visual field.