CSB332H1 Study Guide - Final Guide: Extrapyramidal System, Glucocorticoid, 5-Ht1A Receptor

25 Apr 2014
School
Department
Course
Professor
Document Summary
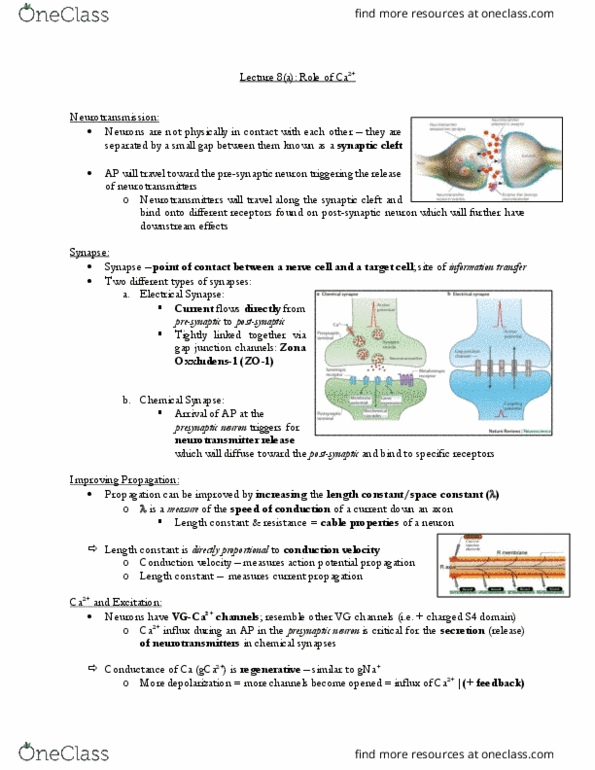
Neurotransmitter is released into the synaptic cleft. Autoreceptors meaning receptors for the self (e. g. , self-receptors). Autoreceptors can be located on the presynaptic axon or somatic dendritic sites of the neuron (e. g. , cell bodies, dendrites). This is another mechanism that regulates neurotransmitter release. Ne binds to the 2-adrenergic receptor on its own presynaptic axon. Autoreceptors are typically coupled to an inhibitory g protein (go or gi). 2- adrenergic receptor is coupled to a gi protein. When activated by ne, the beta and gamma subunits of the g protein will dissociate and will decrease the conductance of the ca2+ channel. It will prevent the further influx of ca2+ into presynaptic axon. The increase in the levels of presynaptic ca2+ is responsible for exocytosis of neurotransmitters. The decrease in ca2+ concentrations would lead to the prevention of further release of ne in synaptic cleft. This is a third mechanism regulating neurotransmitter release.