CSB331H1 Lecture Notes - Lecture 6: Phosphatidylinositol 4,5-Bisphosphate, Brownian Ratchet, Microtubule

4 Apr 2012
School
Department
Course
Professor
Document Summary
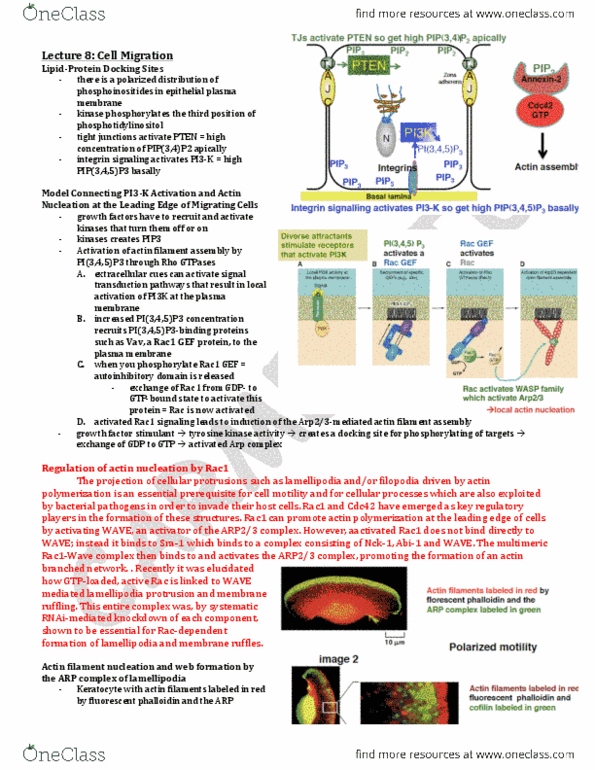
The majority of the notes is verbatim from the 4th and 5th editions of alberts and cited literature. While actin polymerization can push plasma membranes to generate cellular protrusions at the leading edge, movement of cells is also dependent on the interaction of actin cytoskeleton with myosin motors. Myosin motors move along actin filament by hydrolysis of atp to transform chemical energy into mechanical energy- hence myosin motors are often referred to as mechanochemical motors. As noted in alberts, the myosin family is composed of several members. The focused is on myosin i and myosin ii as the play key roles in promoting cellular movements. The tail of myosin i interacts with lipid bilayers, by a still poorly understood mechanism. Via its interactions with the plasma membrane and actin-filaments at the leading edge of migrating cells, it"s hypothesized that myosin i promotes lamellipodia and filopodia extensions.