CSB331H1 Study Guide - Midterm Guide: Pdz Domain, Adherens Junction, Cell Junction

7 Sep 2012
School
Department
Course
Professor


Document Summary
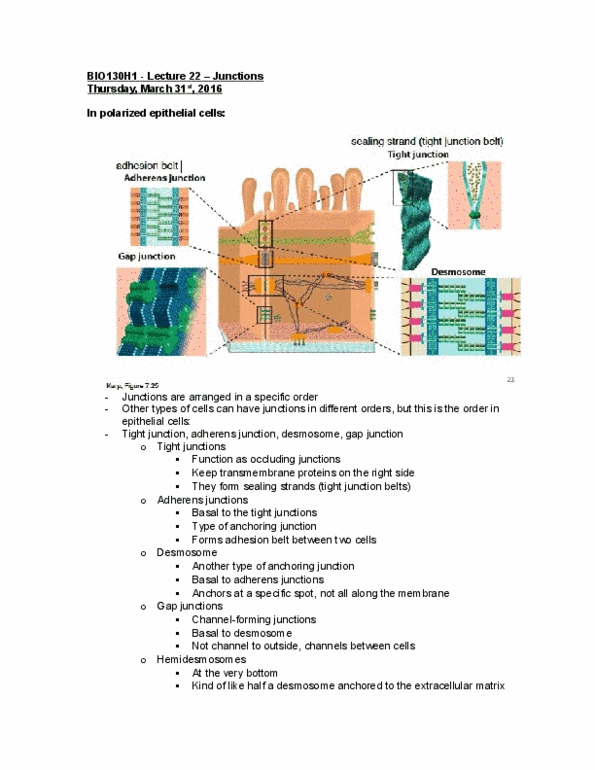
Epithelial tissue mechanical stress transmitted from cell to cell across the whole epithelia by cytoskeletal filaments anchored to cell-matrix and cell-cell adhesion sites. Connective tissue extracellular matrix directly bears mechanical stress of tension and compression. Occluding junctions: tight junctions regulate what can pass through epithelia. Cell-cell anchoring junctions: adherens junctions link actin filaments between cells, desmosomal junctions link intermediate filaments between cells. Integrins anchor actin filaments: hemidesmosomes anchor intermediate filaments. Light microscopy cannot resolve details smaller than its own wavelength. Cell-cell junctions can be visualized with electron microscopy: use biochemical and molecular biology methods to study function. Adherens junctions connects actin filament bundle in one cell with that in the next cell. Cadherin (classical cadherin) is the transmembrane adhesion protein that mediates adherens junctions: e-cadherin (many epithelia, n-cadherin (neurons, heart, skeletal, muscle, fibroblasts, p-cadherin (placenta, epidermis, breast epithelium, ve-cadherin (endothelial cells) E-cadherin expression confers the polarized epithelial phenotype: loss of e-cadherin = emt, gain of e-cadherin = met.