BIOL 112 Lecture Notes - Lecture 11: Glycogen, Ion Channel, Salami

27 Apr 2017
School
Department
Course
Professor


27
BIOL 112 Full Course Notes
Verified Note
27 documents
Document Summary
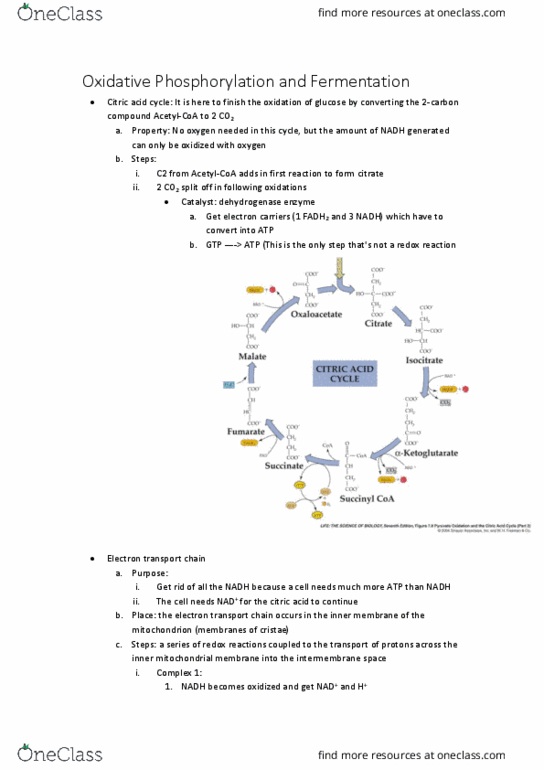
The end product of glycolysis is pyruvate (this occurs in the cytosol). Then, pyruvate is oxidized in the mitochondrial matrix to acetyl-coenzyme a. Acetyl-coa: c-s is a high-energy bond , so acetyl-coa is an energy carrier like atp; it allows transfer of that 2-carbon group to another molecule at the start of the citric acid cycle. This high- energy bond can again be generated by the large amount of free energy released by the pyruvate/nad+ redox reaction. Acetyl-coa is a key molecule in metabolism, because sugars, fats and many amino acids are all broken down to acetyl-coa, and acetyl-coa also serves as the building block for making fats and other metabolites. The pyruvate move to the mitochondria matrix where it gets oxidized and then goes into the citric acid cycle (crebbs cycle) in the mitochondria matrix still. Citric acid cycle: it is there to finish the oxidation of glucose by converting the 2- carbon compound acetyl-coa to 2 co2.